Cuando amanece, en África, la gacela despierta.
Ella sabe que deberá correr más aprisa
que el más veloz de los leones o morirá devorada.
Cuando amanece, en África, el león despierta.
Él sabe que deberá correr más aprisa
que la más lenta de las gacelas o morirá de hambre.
No importa si eres león o gacela, cuando salga el sol, corre
tan rápido como puedas.
(poema africano)
cc |
|
 No
sabemos si en 1927 el joven ecólogo Charles Elton se inspiró
en este poema africano para desarrollar su teoría de las
cadenas tróficas, pero sin duda también el fue capaz
de explicar claramente que todos los seres vivos podemos servir
de comida para otros. No
sabemos si en 1927 el joven ecólogo Charles Elton se inspiró
en este poema africano para desarrollar su teoría de las
cadenas tróficas, pero sin duda también el fue capaz
de explicar claramente que todos los seres vivos podemos servir
de comida para otros.
Elton imaginó la estructura del ecosistema con la forma de
una pirámide: En la base situó a los vegetales y en
secciones cada vez más altas y estrechas a los herbívoros
y después a los carnívoros. A cada uno de estos grupos
los llamó niveles tróficos, (del griego throphos:
alimento) dejando claro que todos los seres vivos nos transformamos
en comida después de un breve paso por la vida, contribuyendo
a mantener en pié la estructura del ecosistema.
Este enfoque descriptivo de la Naturaleza llevó a la comunidad
científica a ensayar todo tipo de cálculos, aplicando
las herramientas matemáticas y las técnicas analíticas
más sofisticadas del momento. Fue en aquellos años
cuando Lotka y Volterra enunciaron su conocido modelo depredador-presa
aplicando las ecuaciones diferenciales para explicar la relación
depredador / presa (Fig.1). También los oceanógrafos
del buque danés Galathea utilizaron la absorción del
C14 para determinar la cantidad de materia orgánica producida
por los vegetales unicelulares del océano por unidad de superficie
y año. A este valor lo llamaron Producción Primaria
Neta y se fijó en un máximo de 100 gr/m2/año.
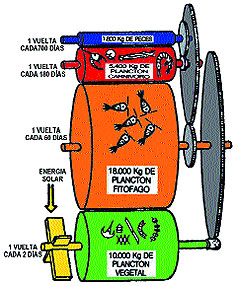
Según estos cálculos, Lindeman enunció su famosa
Ley del 10%: “Tan sólo el 10% de las calorías
ingeridas por un nivel de la pirámide trófica es transferido
al nivel siguiente, el 90% restante es utilizado para mantener la
vida en ese nivel o bien es excretado sin pasar al nivel trófico
siguiente”
Esta escasa eficiciencia en el transporte de la materia orgánica
de un nivel trófico al siguiente limita a 4 el número
máximo de eslabones de una cadena trófica. En otras
palabras, 1.000 gr. de fitoplancton pueden producir 100 gr. de copépodos
herbívoros, que a su vez se transformarán en 10 gr.
de sardina y después en 1 gr de tiburón.
  Todo
parecía encajar bien en la descripción de los ecosistemas
marinos, incluso se demostró triunfalmente que una pequeña
cantidad de biomasa de fitoplancton permitía alimentar una
abundante biomasa del nivel trófico superior (zooplancton
herbívoro) ya que se comprobó experimentalmente su
altísima tasa de renovación. Todo
parecía encajar bien en la descripción de los ecosistemas
marinos, incluso se demostró triunfalmente que una pequeña
cantidad de biomasa de fitoplancton permitía alimentar una
abundante biomasa del nivel trófico superior (zooplancton
herbívoro) ya que se comprobó experimentalmente su
altísima tasa de renovación.
Sin embargo algunas observaciones del mundo real no encajaban con
esta interpretación simplificada de la Naturaleza.
Se descubrió que muchos animales cambian de herbívoro
a carnívoro en su ciclo vital, de modo que aunque en todas
las cadenas hay productores, herbívoros, carnívoros
y carroñeros, no se corresponden con grupos fijos de organismos
puesto que las respectivas funciones pueden ir cambiando a lo largo
de sus ciclos biológicos.
Actualmente se acepta que ningún sistema trófico ha
sido descrito en términos realmente cuantitativos, tal es
la complejidad de lo que ahora llamamos redes tróficas.
Las cadenas alimenticias son en realidad más complejas de
lo que pensábamos, de modo que los eslabones de estas cadenas
suelen estar conectados entre si de diversas maneras para formar
una intrincada y tupida red alimenticia.
La clásica pirámide de Elton ha sido sustituida por
la pirámide energética y la disciplina científica
que aborda estos estudios es ahora la termodinámica de procesos
irreversibles.
 Hoy
sabemos que la sucesión de eslabones tróficos tiene
como objeto casi exclusivo la transferencia de energía y
que, posiblemente, la función del ecosistema sea tan sólo
maximizar la permanencia de esta energía dentro de estructuras
biológicas. Hoy
sabemos que la sucesión de eslabones tróficos tiene
como objeto casi exclusivo la transferencia de energía y
que, posiblemente, la función del ecosistema sea tan sólo
maximizar la permanencia de esta energía dentro de estructuras
biológicas.
Por esta razón todos los seres vivos se resisten a ser devorados
y han desarrollado estrategias de todo tipo para escapar de su depredador
y atesorar la energía del sistema en sus propias estructuras.
Se confirma también que la energía no fluye siempre
de niveles tróficos inferiores a superiores; puede transferirse
también lateralmente, cuando especies en competencia se comen
unas a otras o hasta endógenamente, cuando se da el canibalismo
en ciertas especies.
No conocemos con suficiente detalle las relaciones entre los organismos
marinos como para poder describir con exactitud las transferencias
de energía en el ecosistema, por esta razón nuestra
comprensión de las cadenas alimenticias marinas permanece
aún a un nivel muy elemental.
En el futuro el conocimiento fino de estos procesos tendrá
su aplicación práctica en la gestión pesquera,
de modo que podremos calcular el volumen de extracción de
biomasa marina sin que se produzcan alteraciones ecológicas.
También nos servirá para desarrollar eficazmente la
industria de la acuicultura, obteniendo recursos marinos a base
de acelerar, en entornos confinados, el funcionamiento de cadenas
alimenticias controladas.
|